Entre los distintos tipos de biomoléculas orgánicas que forman parte de las células vivas hay que distinguir por un lado a las proteínas y los ácidos nucleicos, cuya misión fundamental es el almacenamiento, transmisión y expresión de la información genética, y por otro a los glúcidos y lípidos cuya principal misión es la de proporcionar energía para los distintos procesos celulares y que por lo tanto están llamados a ser los grandes protagonistas del catabolismo. De todos modos, dado que las células se encuentran en un continuo proceso de renovación de sus componentes moleculares, el catabolismo comprende rutas que permiten llevar a cabo la degradación de todas y cada una de las biomoléculas. En este capítulo analizaremos las distintas rutas catabólicas con especial incidencia en aquellas responsables de la degradación de glúcidos y lípidos, por ser éstas las que más frecuentemente utilizan las células.
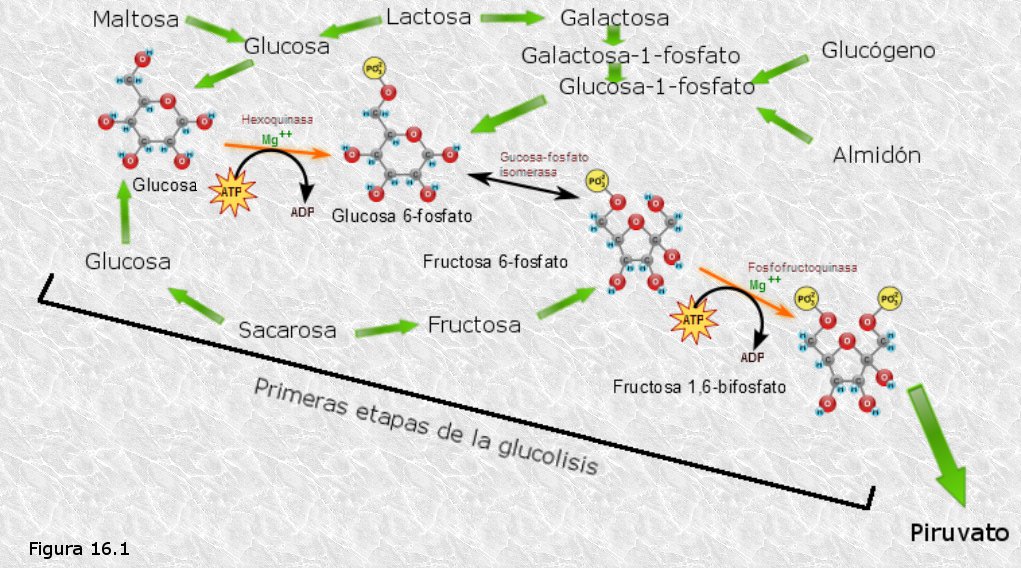
Por ser la glucosa el monosacárido más abundante en la naturaleza, la degradación de los azúcares se lleva a cabo "vía glucosa". Así, el catabolismo de los azúcares converge hacia una única ruta central de degradación de la glucosa.
Los grandes polisacáridos de reserva, mediante una reacción de fosforilación catalizada por la glucógeno-fosforilasao por la almidón fosforilasa según se trate de glucógeno o de almidón, liberan unidades de glucosa-1-fosfato, que a continuación, por acción de la fosfoglucomutasa se transforma en glucosa-6-fosfato, la cual es el primer intermediario de la ruta de degradación de la glucosa.
Los monosacáridos diferentes de la glucosa, que en ocasiones pueden proceder de la hidrólisis de distintos tipos de oligosacáridos, se transforman en glucosa o en algunos de los intermediarios de su degradación, mediante reacciones de isomerización.
Una vez transformados en glucosa los azúcares se pueden degradar completamente hasta CO2 Y H2O siguiendo un camino que incluye tres rutas metabólicas principales: Glucolisis, Ciclo de Krebs y Cadena respiratoria. Existe además una ruta alternativa, la Ruta de las pentosas.
GLUCOLISIS
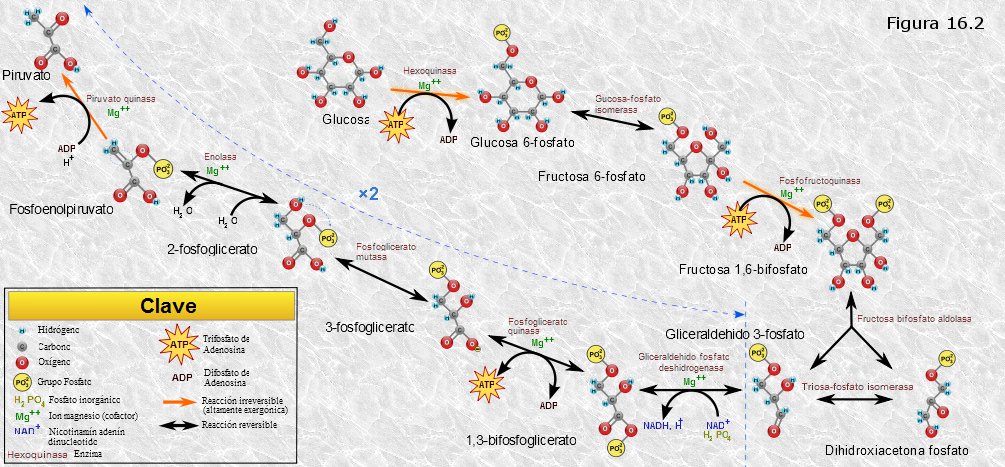
Es un conjunto de reacciones consecutivas que degradan la glucosa , transformándola en dos moléculas de ácido pirúvico. Estas reacciones son anaerobias, es decir, no participa en ellas el oxígeno, y transcurren íntegramente en el hialoplasma celular. Concomitantemente parte de la energía química de la glucosa es recuperada en forma de ATP y NADH.
Probablemente la glucolisis haya sido, en el curso de la evolución biológica, la primera ruta metabólica de la que las células dispusieron para obtener energía. También fue la primera ruta que los bioquímicos conocieron en su totalidad.
Podemos agrupar estas 10 reacciones en dos fases bien diferenciadas:
1.- La glucosa se degrada para dar lugar a dos moléculas de gliceraldehido-3-fosfato. Para ello hace falta energía que es aportada por dos moléculas de ATP.
2.- Las dos moléculas de gliceraldehido-3-fosfato se transforman en dos moléculas de ácido pirúvico. La energía liberada en este proceso es recuperada en forma de cuatro moléculas de ATP y dos de NADH. Es fundamentalmente la oxidación del gliceraldehido-3-fosfato en la reacción 6 la que libera la energía química, que se recupera en parte en forma de NADH y en parte, mediante dos fosforilaciones a nivel de sustrato en las reacciones siguientes, en forma de ATP.
Así pues, la glucolisis parece necesitar de una primera fase, en la cual se consume energía del ATP, para que la energía química de la glucosa pueda ser liberada en la segunda fase obteniéndose una cantidad mayor de ATP y también NADH.
Haciendo un balance de lo ocurrido durante la glucolisis, por cada molécula de glucosa degradada se obtienen 2 moléculas de ácido pirúvico, 2 moléculas de NADH y 2 moléculas de ATP (4 obtenidas en la segunda fase menos 2 consumidas en la primera).
El rendimiento energético en moléculas de ATP de la degradación de la glucosa a ácido pirúvico a través de la glucolisis no es muy elevado. La glucosa contiene en sus enlaces mucha más energía química que la que pudo ser recuperada hasta ahora en forma de 2 moléculas de ATP. Parte de esa energía se habrá perdido en forma de calor en el transcurso de la glucolisis, pero otra parte importante permanece en los enlaces del ácido pirúvico y en el NADH, y sólo podrá ser extraída mediante la oxidación de estos compuestos en las reacciones aerobias de la respiración celular que tienen lugar en la mitocondria, en las cuales se producirá una cantidad mucho mayor de ATP.
Sin embargo, algunas formas de vida muy simples, tales como algunas bacterias y levaduras, pueden sobrevivir holgadamente con esta pequeña cantidad de energía que obtienen de la glucolisis, debiendo resolver entonces algunos problemas que analizaremos a continuación.
FERMENTACIONES.
Por cada molécula de glucosa que se degrada en la glucolisis 2 moléculas de NAD+ se transforman en NADH. Puesto que la cantidad de NAD+ que poseen las células es limitada debe existir algún mecanismo que permita oxidar el NADHtransformándolo de nuevo en NAD+ para que pueda ser reutilizado en la glucolisis, de lo contrario todo el proceso se detendría.
En las células aerobias el NADH se oxida a NAD+ cediendo sus electrones al oxígeno (aceptor último de electrones en este tipo de células) a través de la cadena de transporte electrónico mitocondrial.
En las células anaerobias, que no pueden utilizar el oxígeno como aceptor último de electrones en sus oxidaciones metabólicas, el NADH se oxida a NAD+ cediendo sus electrones a una molécula orgánica aceptora, que puede ser el ácido pirúvico o algún producto de la transformación de éste, a través de una o más reacciones adicionales a la glucolisis. Estas etapas adicionales que tienen como único objeto reciclar el NADH obtenido en la glucolisis a NAD+ reciben el nombre defermentaciones.
Las fermentaciones consisten, pues, en la transformación del ácido pirúvico que se obtiene al final de la glucolisis, en algún otro producto orgánico sencillo que es diferente en cada tipo de fermentación. Existen muchos tipos de fermentación que dan lugar a toda una gama de productos, algunos de ellos de interés alimentario o industrial, pero los dos tipos principales son la fermentación homoláctica y la fermentación alcohólica.
a) Fermentación homoláctica.- Tiene lugar en una sola etapa: el ácido pirúvico acepta un par de electrones procedente del NADH con lo cual se reduce a ácido láctico, que es el producto final.
b) Fermentación alcohólica.- Tiene lugar en dos etapas. En la primera el ácido pirúvico pierde su grupo carboxilo en forma de CO2 para dar acetaldehido. En la segunda el acetaldehido acepta un par de electrones procedentes del NADH transformándose en alcohol etílico. Alcohol etílico y CO2 son los productos finales.
La fermentación heteroláctica es un tipo mixto en el que se obtiene una molécula de ácido láctico, una de alcohol etílico y una de CO2.
Existen células, llamadas anaerobias estrictas, que en ningún caso pueden utilizar el oxígeno como aceptor de electrones. En ellas la glucolisis, complementada por algún tipo de fermentación, es la única ruta disponible para la obtención de energía. Dado que cuando la vida se originó sobre la Tierra la atmósfera carecía de oxígeno, es de suponer que las primeras células vivas pertenecían a esta categoría. Prueba de ello es que la mayoría de las células actuales, incluidas las aerobias, conservan la capacidad de fermentar el ácido pirúvico a ácido láctico y la utilizan durante cortos períodos en los que el oxígeno no está disponible, obteniendo entonces energía exclusivamente a partir de la glucolisis. Así ocurre en las células del músculo esquelético, que aunque normalmente trabajan en condiciones aerobias, pueden obtener energía por este procedimiento en ocasiones, como por ejemplo durante un ejercicio muscular intenso, en las que el oxígeno aportado por la sangre resulta insuficiente. En la actualidad es práctica habitual en algunos deportes de alta competición medir de manera periódica durante los entrenamientos los niveles de ácido láctico en sangre de los deportistas, con el objeto de determinar en cada caso el umbral de esfuerzo a partir del cual el músculo comienza a trabajar en anaerobiosis y, en consecuencia, a disminuir su rendimiento.
1.3.-RUTA DE LAS PENTOSAS.
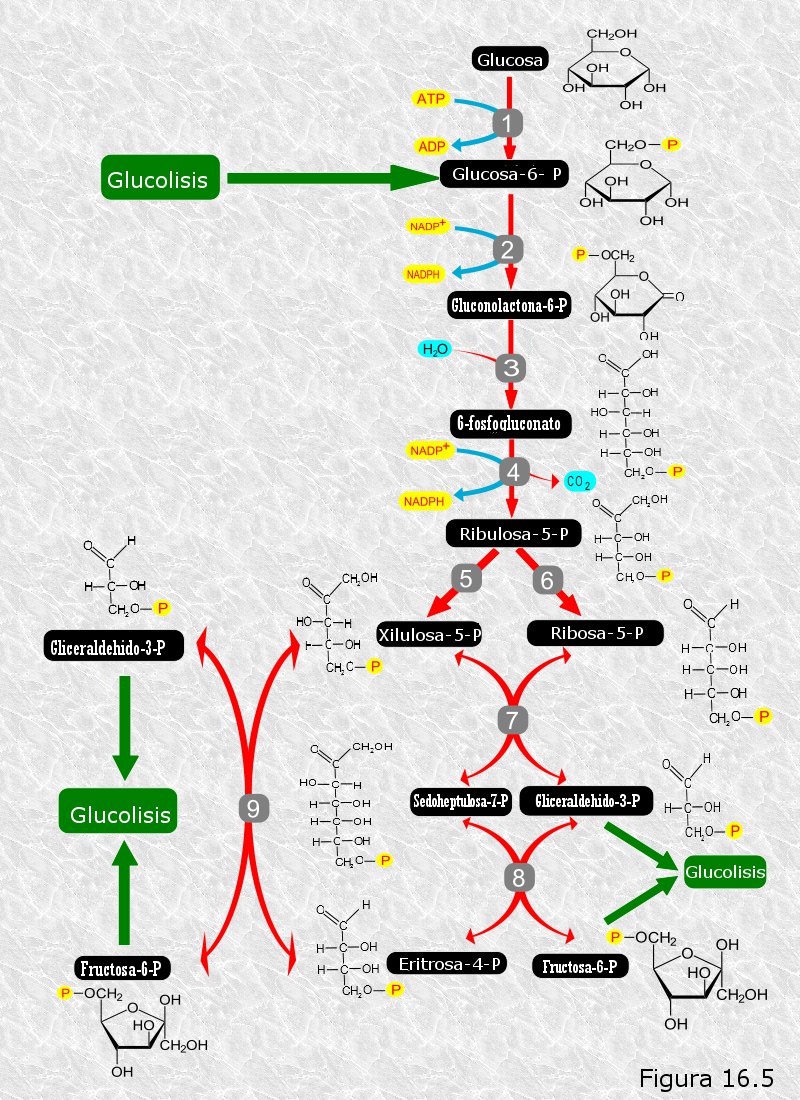
La glucolisis tiene como misión recuperar parte de la energía química de la glucosa en forma de ATP. Sin embargo, las células pueden necesitar en ocasiones degradar la glucosa con otros fines.
La ruta de las pentosas es una vía alternativa a la glucolisis que tiene como finalidad la obtención de azúcares de 5 átomos de carbono, como la ribosa, necesarios para la biosíntesis de nucleótidos, y de coenzima reducidoNADPH necesario para diversos tipos de biosíntesis en el anabolismo.
En la ruta de las pentosas la glucosa-6-fosfato da lugar, mediante dos deshidrogenaciones y una descarboxilación en las que se obtiene NADPH, a una molécula de CO2 y a un azúcar de 5 átomos de carbono, la ribulosa-5-fosfato, que después se transforma en ribosa-5-fosfato.
Dado que los productos de esta ruta se emplean en procesos biosintéticos del anabolismo, podemos considerar que la ruta de las pentosas es una ruta típicamente anfibólica.
OXIDACIONES RESPIRATORIAS.
Las células aerobias pueden oxidar el ácido pirúvico obtenido en la glucolisis en presencia de oxígeno molecular y obtener así una gran cantidad de energía en forma de ATP. El proceso completo de degradación de la glucosa en presencia de oxígeno se conoce como respiración celular.
Desde los tiempos del químico Lavoisier es sabido que el proceso respiratorio consiste en una reacción de la materia orgánica con el oxígeno para dar como productos dióxido de carbono y agua. Esta reacción es similar a la que se produce en la combustión de cualquier tipo de materia orgánica. Se suele decir que las oxidaciones respiratorias son combustiones a baja temperatura o "combustiones sin llama". El proceso de oxidación de la materia orgánica en presencia de oxígeno es fuertemente exergónico y parte de la energía química que en él se libera es recuperada en forma de ATP. La ecuación global de la respiración celular puede escribirse como sigue:
MATERIA ORGÁNICA + O2 → CO2 + H2O
Las reacciones aerobias de la respiración celular tienen lugar íntegramente en la mitocondria, más concretamente en la matriz mitocondrial y en la membrana mitocondrial interna. Veremos más adelante que estas oxidaciones respiratorias no son exclusivas del catabolismo de los azúcares, sino que en gran medida son comunes al catabolismo de otras biomoléculas como lípidos y proteínas. Sin embargo, vamos a centrarnos ahora en los procesos que conducen a la degradación total hasta CO2 y H2O del ácido pirúvico procedente de la glucolisis, a saber, la descarboxilación oxidativa del ácido pirúvico, elciclo de Krebs, y la cadena respiratoria.
DESCARBOXILACIÓN OXIDATIVA DEL ÁCIDO PIRÚVICO.
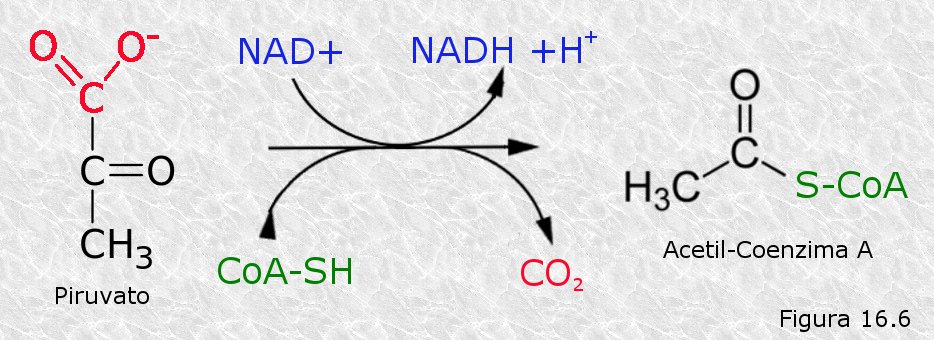
El ácido pirúvico procedente de la glucolisis penetra en la matriz mitocondrial a través de permeasas específicas de las membranas mitocondriales. Una vez allí, sufre una descarboxilación oxidativa catalizada por el complejo de la piruvato deshidrogenasa, dando lugar a una molécula de CO2 y a un grupo acetilo del acetil-coenzima A (Figura 16.6). Por cada molécula de ácido pirúvico se consume una de coenzima A (un coenzima transportador de grupos acilo) y una molécula deNAD+ es reducida a NADH. Hay que destacar que, dado que cada molécula de glucosa da lugar a dos de ácido pirúvico en la glucolisis, y que en la descarboxilación de cada una de éstas se desprende una molécula de CO2, dos de los seis átomos de carbono de la glucosa se han desprendido ya en forma de estas dos moléculas de CO2, que es uno de los productos finales de la respiración celular.
CICLO DE KREBS.
El acetil-CoA obtenido en la etapa anterior puede ser ahora oxidado en la misma matriz mitocondrial mediante una ruta metabólica cíclica llamada ciclo de los ácidos tricarboxílicos o ciclo del ácido cítrico, y que, en honor a su descubridor, es más conocida por ciclo de Krebs.
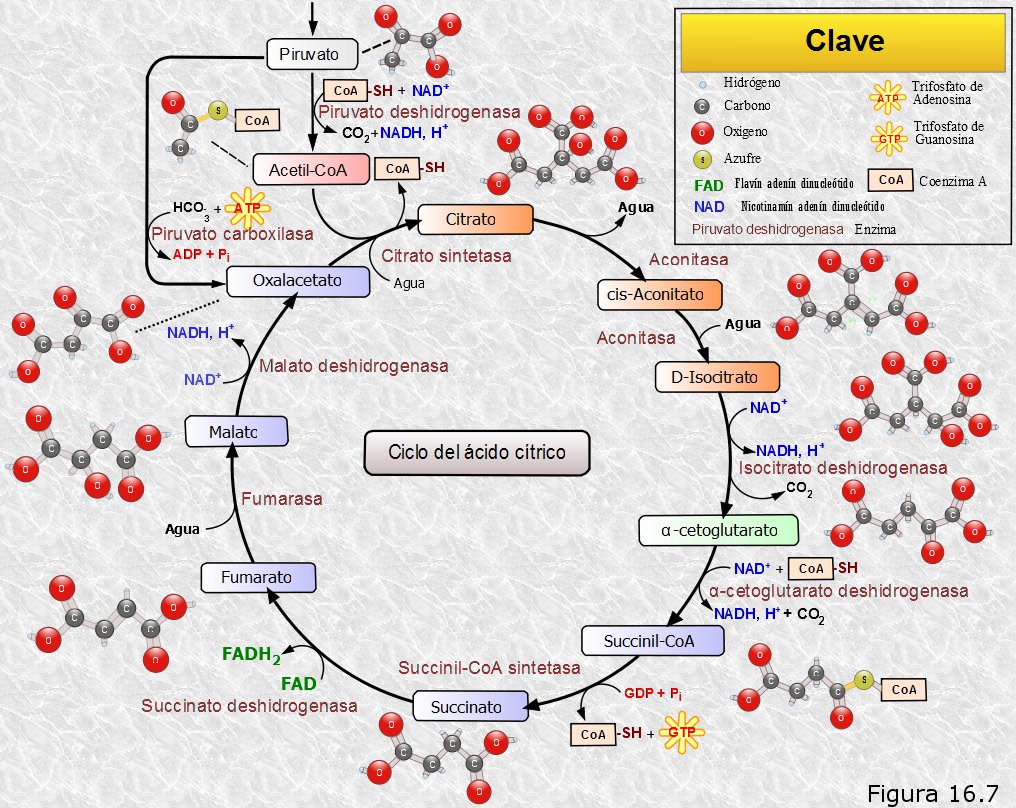
El ciclo se inicia con la condensación del grupo acetilo del acetil-CoA (2C) con una molécula ácido oxalacético (4C) para dar un ácido tricarboxílico de 6 átomos de carbono, el ácido cítrico. En esta reacción se libera el coenzima A. Posteriormente, en una secuencia de siete reacciones catalizadas enzimáticamente, se eliminan dos átomos de carbono en forma de CO2 y se regenera el ácido oxalacético.
A lo largo del ciclo se producen dos descarboxilaciones oxidativas, en las etapas 3 y 4, que liberan dos moléculas de CO2. Con ello se completa la degradación del esqueleto carbonado de la glucosa: dos átomos de carbono se desprendieron en
forma de dos moléculas de CO2 en la descarboxilación oxidativa de dos moléculas de ácido pirúvico; las dos moléculas de acetil-CoA resultantes ingresan en el ciclo de Krebs liberándose por cada una de ellas otras dos moléculas de CO2; en total se han liberado seis moléculas de CO2 que se corresponden con los seis átomos de carbono de la glucosa.
Por otra parte, las distintas oxidaciones que tienen lugar a lo largo del ciclo liberan electrones, que en unos casos son captados por el NAD+ (reacciones 3, 4 y 8) y en otros por el FAD (reacción 6), los cuales se reducen respectivamente aNADH y FADH2.
Además, en la reacción 5, tiene lugar una fosforilación a nivel de sustrato, consecuencia de la anterior oxidación, que da lugar a una molécula de GTP, posteriormente transformable en ATP.
Haciendo un balance de lo ocurrido en el ciclo de Krebs, por cada grupo acetilo del acetil-CoA que ingresa en el mismo se obtienen 2 moléculas de CO2, 3 moléculas de NADH, 1 molécula de FADH2 y una molécula de GTP transformable en ATP.
El rendimiento energético en moléculas de ATP obtenido hasta el momento no parece muy alto: sólo dos moléculas deATP en la glucolisis y otras dos en el ciclo de Krebs (una por cada acetil-CoA). Sin embargo, una gran parte de la energía química de la glucosa está temporalmente retenida en forma de "electrones de alta energía" de los coenzimas reducidosNADH y FADH2 que se fueron obteniendo a lo largo de todo el proceso. Será en la siguiente etapa de las oxidaciones respiratorias cuando estos coenzimas cedan sus electrones al oxígeno, produciéndose así, por fin, la liberación de toda esa energía, una parte de la cual será recuperada en forma de ATP

 En función del tamaño y la naturaleza de las partículas ingeridas, la endocitosis puede ser de dos tipos: pinocitosis y endocitosis. Pinocitosis: Es la ingestión de pequeñas partículas o líquidos, mediante la formación de vesículas muy pequeñas. Se da en todo tipo de células. Fagocitosis: consiste en la ingestión de partículas de gran tamaño, organismos vivos o restos celulares que forman unas vesículas llamadas vesículas o vacuolas de fadocitosis (fagosomas). Este proceso es característico de ciertas células del sistema inmunitario, como los macrófagos y neutrófilos que ingieren partículas extrañas. Exocitosis: Es la secreción de macromoléculas y partículas hacia el medio externo de la célula. Al igual que la endocitosis, implica la fusión con la membrana plasmática de vesículas procedentes del citoplasma celular. Los fenómenos de exocitosis desempeñan un papel importante en diversas funciones celulares: Funciones estructurales y de relación: consiste en la secreción de sustancias sintetizadas en el interior de la célula y cuyo destino final es, por ejemplo, la formación del glicocáliz o la matriz celular, o bien el intercambio de metabolitos o señales con otras células. La secreción puede ser de dos tipos: -. Constitutiva: se realiza de forma continua a partir de vesículas originadas en el sistema retículo endoplasmático-Golgi y guarda relación con sustancias que van a tener una función estructural, como, por ejemplo, la renovación de la membrana o el glicocáliz. -. Regulada: se produce en lugares localizados de la célula ante determinados estímulos externos. Es típica de células secretoras de las glándulas exocrinas (secreción de enzimas digestivas) o endocrinas (secreción de hormonas), y también de la liberación neuronal de neurotransmisores. En este caso, como en el de la endocitosis mediada por receptor, las vesículas también están revestidas de clatrina. Funciones de excreción: se trata de la secreción de los productos de desecho que se producen tras la digestión celular de partículas ingeridas por fagocitosis.
En función del tamaño y la naturaleza de las partículas ingeridas, la endocitosis puede ser de dos tipos: pinocitosis y endocitosis. Pinocitosis: Es la ingestión de pequeñas partículas o líquidos, mediante la formación de vesículas muy pequeñas. Se da en todo tipo de células. Fagocitosis: consiste en la ingestión de partículas de gran tamaño, organismos vivos o restos celulares que forman unas vesículas llamadas vesículas o vacuolas de fadocitosis (fagosomas). Este proceso es característico de ciertas células del sistema inmunitario, como los macrófagos y neutrófilos que ingieren partículas extrañas. Exocitosis: Es la secreción de macromoléculas y partículas hacia el medio externo de la célula. Al igual que la endocitosis, implica la fusión con la membrana plasmática de vesículas procedentes del citoplasma celular. Los fenómenos de exocitosis desempeñan un papel importante en diversas funciones celulares: Funciones estructurales y de relación: consiste en la secreción de sustancias sintetizadas en el interior de la célula y cuyo destino final es, por ejemplo, la formación del glicocáliz o la matriz celular, o bien el intercambio de metabolitos o señales con otras células. La secreción puede ser de dos tipos: -. Constitutiva: se realiza de forma continua a partir de vesículas originadas en el sistema retículo endoplasmático-Golgi y guarda relación con sustancias que van a tener una función estructural, como, por ejemplo, la renovación de la membrana o el glicocáliz. -. Regulada: se produce en lugares localizados de la célula ante determinados estímulos externos. Es típica de células secretoras de las glándulas exocrinas (secreción de enzimas digestivas) o endocrinas (secreción de hormonas), y también de la liberación neuronal de neurotransmisores. En este caso, como en el de la endocitosis mediada por receptor, las vesículas también están revestidas de clatrina. Funciones de excreción: se trata de la secreción de los productos de desecho que se producen tras la digestión celular de partículas ingeridas por fagocitosis.